This readme file is for the model published in:
Grunditz A, Holbro N, Tian L, Zuo Y and Oertner TG
Spine neck plasticity controls postsynaptic calcium signals through
electrical compartmentalization
J Neuroscience (2008) 28: 13457-13466
This model was set up to dissect the relative contribution of
different channels to the spine calcium transients measured at single
spines. Our model spine is equipped with NMDA and AMPA receptors and
R-type calium channels. Channel densities and spine neck resistance
(Rneck) were adjusted to reproduce the relative amplitude of
fluorescence transients measured in our pharmacological experiments
How to run the model Under Windows:
1)download archive from ModelDB and expand the zip file
2) compile the mod files with mknrndll
3) start the program by double clicking the mosinit.hoc file
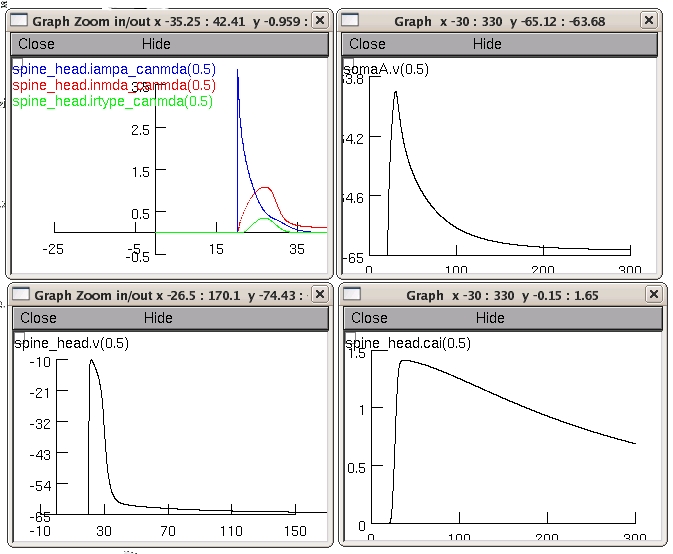
then you will hopefully see a 4 graphs plotting:
1) the EPSP at the soma
2) the EPSP at the spine head
3) the concentration calcium (bound to dye molecules) in the spine
head
4) the current though the three channels (AMPARs, NMDARs and R-type
channels) similar to the simulations in figure 4 of the paper
 Comments:
Morphology of our CA1 pyramidal cell: we used the multi-compartment
model of a CA1 pyramidal cell from the NEURON database published in
Golding NL, Kath WL, Spruston N
Dichotomy of action-potential backpropagation in CA1 pyramidal neuron
dendrites
J Neurophysiol (2001) 86: 2998-3010.
The model was simplified by removing all active conductances from the
soma and dendrites. We set membrane time constant (tm) to 28 ms,
giving Rm = 28 kOcm2 and Cm = 1uF/cm2. An intracellular resistivity of
Ri = 150 Ocm , and a membrane resting potential of Vrest = -65 mV were
used.
Three types of channels were implemented in the spine compartment,
namely NMDA, AMPA, and R-type voltage gated calcium channels.
The voltage-dependent R-type calcium conductance was simulated using
Hodgkin-Huxley like equation adapted from
Foehring RC, Mermelstein PG, Song WJ, Ulrich S, Surmeier DJ Unique
properties of R-type calcium currents in neocortical and neostriatal
neurons J Neurophysiol (2000) 84:2225-2236.
The kinetic equations for the AMPA and NMDA mechanism were taken from
Franks KM, Bartol TM, Jr., Sejnowski TJ A Monte Carlo model reveals
independent signaling at central glutamatergic synapses Biophysical
Journal 2002 83:2333-2348.
Note: The NMDA current was calculated in three steps:
First as an non specific ion current though the NMDA receptor
Second the calcium current through the NMDA receptors was calculated
Third a balance current thorough the NMDA receptors was substracted
from the total current through the NMDA receptors
To model accumulation and diffusion of calcium-bound dye, we used a
mechanism taken from the NERUON database, that simulates radial
diffusion between concentric shells inside a compartment and
longitudinal diffusion between adjacent compartments. The diffusion
mechanism was inserted to the spine, spine neck, and connected
dendritic compartments (D = 0.23 um2/ms, diffusion coefficient of
fluo5F-Ca2+ in cytoplasm).
Questions on how to use this model should be directed to
asa.mueller@fmi.ch
Comments:
Morphology of our CA1 pyramidal cell: we used the multi-compartment
model of a CA1 pyramidal cell from the NEURON database published in
Golding NL, Kath WL, Spruston N
Dichotomy of action-potential backpropagation in CA1 pyramidal neuron
dendrites
J Neurophysiol (2001) 86: 2998-3010.
The model was simplified by removing all active conductances from the
soma and dendrites. We set membrane time constant (tm) to 28 ms,
giving Rm = 28 kOcm2 and Cm = 1uF/cm2. An intracellular resistivity of
Ri = 150 Ocm , and a membrane resting potential of Vrest = -65 mV were
used.
Three types of channels were implemented in the spine compartment,
namely NMDA, AMPA, and R-type voltage gated calcium channels.
The voltage-dependent R-type calcium conductance was simulated using
Hodgkin-Huxley like equation adapted from
Foehring RC, Mermelstein PG, Song WJ, Ulrich S, Surmeier DJ Unique
properties of R-type calcium currents in neocortical and neostriatal
neurons J Neurophysiol (2000) 84:2225-2236.
The kinetic equations for the AMPA and NMDA mechanism were taken from
Franks KM, Bartol TM, Jr., Sejnowski TJ A Monte Carlo model reveals
independent signaling at central glutamatergic synapses Biophysical
Journal 2002 83:2333-2348.
Note: The NMDA current was calculated in three steps:
First as an non specific ion current though the NMDA receptor
Second the calcium current through the NMDA receptors was calculated
Third a balance current thorough the NMDA receptors was substracted
from the total current through the NMDA receptors
To model accumulation and diffusion of calcium-bound dye, we used a
mechanism taken from the NERUON database, that simulates radial
diffusion between concentric shells inside a compartment and
longitudinal diffusion between adjacent compartments. The diffusion
mechanism was inserted to the spine, spine neck, and connected
dendritic compartments (D = 0.23 um2/ms, diffusion coefficient of
fluo5F-Ca2+ in cytoplasm).
Questions on how to use this model should be directed to
asa.mueller@fmi.ch